ESTRUCTURA Y FUNCIÓN CELULAR
CLASE 1: BIOMOLÉCULAS O BIOCOMPUESTOS
GLÚCIDOS
Estructura química:
Tipos de glúcidos:
Los glúcidos se dividen en monosacáridos, disacáridos u oligosacáridos y polisacáridos.
Para entender la vida tal como la conocemos, primero debemos entender un poco de química orgánica. Las moléculas orgánicas contienen carbono e hidrógeno básicamente. Mientras que muchos químicos orgánicos también contienen otros elementos, es la unión del carbono - hidrógeno lo que los define como orgánicos.
Los compuestos que forman parte esencial de los seres vivos son denominados biocompuestos. Los biocompuestos fundamentalmente se conforman del elemento carbono y entre su clasificación se encuentran los carbohidratos o azúcares, los lípidos o grasas, las proteínas y las vitaminas, entre otros.
Algunas de esta moléculas, como los hidratos de carbono, las proteínas y los ácidos nucléicos pueden ser poliméricas. Se denomina polímero a toda macromolécula constituida por la unión de muchas moléculas pequeñas similares, las que reciben el nombre de monómeros. Cuando dos monómeros similares se unen forman un dímero, si son tres un trímero. Hasta diez se lo nombran genéricamente oligómero.
Los glúcidos, carbohidratos o sacáridos (del griego σάκχαρον que significa "azúcar") son una clase de biomoléculas. Son la forma biológica primaria de almacenamiento y consumo de energía. Otras moléculas que también aportan energía son son las grasas y, en menor medida, las proteínas.
El término hidrato de carbono o carbohidrato es poco apropiado, ya que estas moléculas no son átomos de carbono hidratados, es decir, enlazados a moléculas de agua, sino de átomos de carbono unidos a otros grupos funcionales químicos. Este nombre proviene de la nomenclatura química del siglo XIX, ya que las primeras sustancias aisladas respondían a la fórmula elemental Cn(H2O)n (donde "n" es un entero=1,2,3... según el número de átomos). De aquí el término "carbono-hidratado" se haya mantenido, si bien posteriormente se vio que otras moléculas con las mismas características químicas no se corresponden con esta fórmula. Además, los textos científicos anglosajones aún insisten en denominarlos carbohydrates lo que induce a pensar que este es su nombre correcto. Del mismo modo, en dietética, se usa con más frecuencia la denominación de carbohidratos.
Estructura química:
Los glúcidos son compuestos formados en su mayor parte por átomos de carbono e hidrógeno y en una menor cantidad de oxígeno. Los glúcidos tienen enlaces químicos difíciles de romper llamados covalentes, mismos que poseen gran cantidad de energía, que es liberada al romperse estos enlaces. Una parte de esta energía es aprovechada por el organismo consumidor, y otra parte es almacenada en el organismo.
En la naturaleza se encuentran en los seres vivos, formando parte de biomoléculas aisladas o asociadas a otras como las proteínas y los lípidos.
Los glúcidos se dividen en monosacáridos, disacáridos u oligosacáridos y polisacáridos.
Monosacáridos:
Los glúcidos más simples, los monosacáridos, están formados por una sola molécula; no pueden ser hidrolizados a glúcidos más pequeños. La fórmula química general de un monosacárido no modificado es (CH2O)n, donde n es cualquier número igual o mayor a tres. Los monosacáridos poseen siempre un grupo carbonilo en uno de sus átomos de carbono y grupos hidroxilo en el resto, por lo que pueden considerarse polialcoholes.
Los monosacáridos se clasifican de acuerdo a tres características diferentes: la posición del grupo carbonilo, el número de átomos de carbono que contiene y su quiralidad. Si el grupo carbonilo es un aldehido, el monosacárido es una aldosa; si el grupo carbonilo es una cetona, el monosacárido es una cetosa. Los monosacáridos más pequeños son los que poseen tres átomos de carbono, y son llamados triosas; aquéllos con cuatro son llamados tetrosas, lo que poseen cinco son llamados pentosas, seis son llamados hexosas y así sucesivamente. Los sistemas de clasificación son frecuentemente combinados; por ejemplo, la glucosa es una aldohexosa (un aldehido de seis átomos de carbono), la ribosa es una aldopentosa (un aldehido de cinco átomos de carbono) y la fructosa es una cetohexosa (una cetona de seis átomos de carbono).

Ejemplo de un monosacárido de 3 átomos de carbono (Gliceraldehído)
Ciclación:
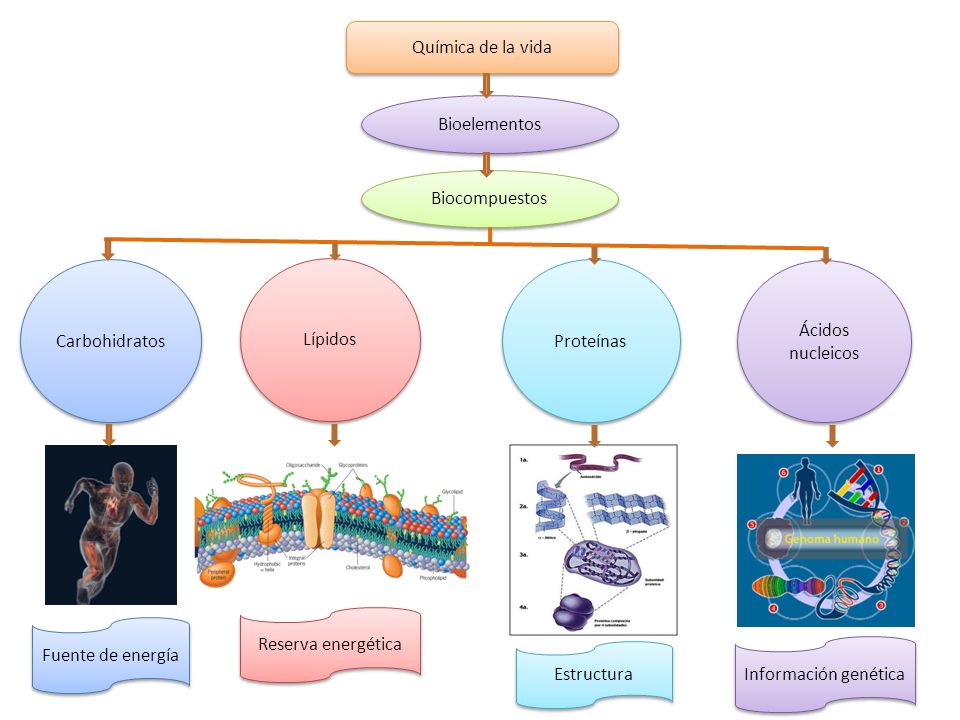
El grupo aldehido o cetona en una cadena lineal abierta de un monosacárido reaccionará reversiblemente con el grupo hidroxilo sobre un átomo de carbono diferente en la misma molécula para formar un hemiacetal o hemicetal, formando un anillo heterocíclico, con un puente de oxígeno entre los dos átomos de carbono. Los anillos con cinco y seis átomos son llamados formas furanosa y piranosa respectivamente y existen en equilibrio con la cadena lineal abierta.

Durante la conversión de la forma lineal abierta a la forma cíclica, el átomo de carbono conteniendo el oxígeno carbonilo, llamado el carbono anomérico, se transforma en un centro quiral con dos posibles configuraciones: el átomo de oxígeno puede tomar una posición arriba o abajo del plano del anillo. El par de estereoisómeros resultantes son llamados anómeros. En el α-anómero, el -OH sustituyente sobre el carbono anomérico sin cuenta en el lado opuesto del anillo (posición trans) a la cadena CH2OH. La forma alternativa, en la cual el sustituyente CH2OH y el grupo hidroxilo sobre el carbono anomérico están en el mismo lado (posición cis) del plano del anillo, es llamado β-anómero. Como el anillo y la forma abierta se interconvierten, ambos anómeros existen en equilibrio.
Los monosacáridos son la principal fuente de combustible para el metabolismo, siendo usado tanto como una fuente de energía (la glucosa es la más importante en la naturaleza) y en biosíntesis. Cuando los monosacáridos no son necesitados para las células son rápidamente convertidos en otra forma, tales como los polisacáridos.
Disacáridos:
Los disacáridos son glúcidos formados por dos moléculas de monosacáridos, por tanto, al hidrolizarse producen dos monosacáridos libres. Los dos monosacáridos se unen mediante un enlace covalente conocido como enlace glucosídico, formado vía una reacción de deshidratación, resultando en la pérdida de un átomo de hidrógeno a partir de un monosacárido y un grupo hidroxilo del otro monosacárido, con la consecuente formación de una molécula de H2O, de manera que la fórmula de los disacáridos no modificados es C12H22O11.

Formación del enlace glucosídico
La sacarosa es el disacárido más abundante y la principal forma en la cual los glúcidos son transportados en las plantas. Está compuesto de una molécula de glucosa y una molécula de fructosa.
La lactosa, un disacárido compuesto por una molécula de galactosa y una molécula de glucosa, estará presente naturalmente sólo en la leche. Otro disacárido notable incluyen la maltosa (dos glucosa enlazadas α-1,4) y la celobiosa (dos glucosa enlazadas β-1,4).

Molécula de sacarosa (ejemplo de disacárido)
Los oligosacáridos están compuestos por entre tres y nueve moléculas de monosacáridos que al hidrolizarse se liberan. No obstante, la definición de cuan largo debe ser un glúcido para ser considerado oligo o polisacárido varía según los autores. Según el número de monosacáridos de la cadena se tienen los trisacáridos (como la rafinosa ), tetrasacárido (estaquiosa), pentasacáridos, etc. Los oligosacáridos se encuentran con frecuencia unidos a proteínas, formando las glucoproteínas, como una forma común de modificación tras la síntesis proteica.
Polisacáridos:
Los polisacáridos son cadenas, ramificadas o no, de más de diez monosacáridos. Los polisacáridos representan una clase importantes de polímeros biológicos. Su función en los organismos vivos está relacionada usualmente con estructura o almacenamiento. El almidón es usado como una forma de almacenar monosacáridos en las plantas, siendo encontrado en la forma de amilosa y la amilopectina (ramificada). En animales, se usa el glucógeno en vez de almidón el cual es estructuralmente similar pero más densamente ramificado. Las propiedades del glucógeno le permiten ser metabolizado más rápidamente, lo cual se ajusta a la vida activa de los animales con locomoción.

La celulosa y la quitina son ejemplos de polisacáridos estructurales. La celulosa y es usada en la pared celular de plantas y otros organismos y es la molécula más abundante sobre la tierra. La quitina tiene una estructura similar a la celulosa pero tiene nitrógeno en sus ramas incrementando así su fuerza. Se encuentra en los exoesqueletos de los artrópodos y en las paredes celulares de muchos hongos. Tiene diversos de usos, por ejemplo en hilos para sutura quirúrgica. Otros polisacáridos incluyen la callosa, el xilano y la galactomanosa.

Los glúcidos desempeñan diversas funciones, siendo la de reserva energética y formación de estructuras las dos más importantes. Así, la glucosa aporta energía inmediata a los organismos, y es la responsable de mantener la actividad de los músculos, la temperatura corporal, la tensión arterial, el correcto funcionamiento del intestino y la actividad de las neuronas. La ribosa y la desoxirribosa son constituyentes básicos de los ácidos nucleótidos, tales como el ARN y el ADN.
LÍPIDOS


Ácidos grasos:

Triglecéridos:

Grasas y aceites:
Fosfolípidos:
![[clip_image002%255B4%255D.gif]](https://blogger.googleusercontent.com/img/b/R29vZ2xl/AVvXsEhtWpuThJPuLTt_4gzmA5iEIjLdI3DMfWJbU1cgcJFzewmBgwds_xGi1osUeIuMDIkZ_HRrMBSmbGn7wIc2wEWdaxtPSzjEDEwAcKttg8FM9_bcsUTvmTss1hVZnpGBi1TvajU1bWh44Xo/s400/clip_image002%25255B4%25255D.gif)
Los fosfolípidos son anfipáticos, esto es que son simultaneamente hidrofílicos e hidrofóbicos. La "cabeza" de un fosfolípido es un grupo fosfato cargado negativamente y las dos "colas" son cadenas hidrocarbonadas fuertemente hidrofóbicas.

Denominamos lípidos a un conjunto muy heterogéneo de biomoléculas cuya característica distintiva aunque no exclusiva ni general es la insolubilidad en agua, siendo por el contrario, solubles en disolventes orgánicos (benceno, cloroformo, éter, hexano, etc.). Están constituidas básicamente por tres elementos: carbono (C), hidrógeno (H) y oxígeno (O); en menor grado aparecen también en ellos nitrógeno (N), fósforo (P) y azufre (S).
Los lípidos pueden encontrarse unidos covalentemente con otras biomoléculas como en el caso de los glicolípidos (presentes en las membranas biológicas). También son numerosas las asociaciones no covalentes de los lípidos con otras biomoléculas, como en el caso de las lipoproteínas y de las estructuras de membrana.
Una característica básica de los lípidos, y de la que derivan sus principales propiedades biológicas es la hidrofobicidad. La baja solubilidad de los lipídos se debe a que su estructura química es fundamentalmente hidrocarbonada (alifática, alicíclica o aromática), con gran cantidad de enlaces C-H y C-C. La naturaleza de estos enlaces es 100% covalente y su momento dipolar es mínimo. El agua, al ser una molécula muy polar, con gran facilidad para formar puentes de hidrógeno, no es capaz de interaccionar con estas moléculas. En presencia de moléculas lipídicas, el agua adopta en torno a ellas una estructura muy ordenada que maximiza las interacciones entre las propias moléculas de agua, forzando a la molécula hidrofóbica al interior de una estructura en forma de jaula, que también reduce la movilidad del lípido.
Un ácido graso es una biomolécula de naturaleza lipídica formada por una larga cadena hidrocarbonada lineal, de diferente longitud o número de átomos de carbono, en cuyo extremo hay un grupo carboxilo (son ácidos orgánicos de cadena larga). Cada átomo de carbono se une al siguiente y al precedente por medio de un enlace covalente sencillo o doble. Al átomo de su extremo le quedan libres tres enlaces que son ocupados por átomos de hidrógeno (H3C-). Los demás átomos tienen libres los dos enlaces, que son ocupados igualmente por átomos de hidrógeno ( ... -CH2-CH2-CH2- ...). En el otro extremo de la molécula se encuentra el grupo carboxilo (-COOH) que es el que se combina con uno de los grupos hidroxilos (-OH) de la glicerina o propanotriol, reaccionando con él. El grupo carboxilo tiene carácter ácido y el grupo hidroxilo tiene carácter básico (o alcalino).
En general, se puede formular un ácido graso genérico como R-COOH, donde R es la cadena hidrocarbonada que identifica al ácido en particular.
Los ácidos grasos forman parte de los fosfolípidos y glucolípidos, moléculas que constituyen la bicapa lipídica de todas las membranas celulares. En los mamíferos, incluido el ser humano, la mayoría de los ácidos grasos se encuentran en forma de triglicéridos, moléculas donde los extremos carboxílico (-COOH) de tres ácidos grasos se esterifican con cada uno de los grupos hidroxilos (-OH) del glicerol (glicerina, propanotriol); los triglicéridos (grasas) se almacenan en el tejido adiposo .
Triglecéridos:
El glicerol es un alcohol de tres carbonos, en cada uno de ellos posee un grupo oxidrilo (OH). Cada OH se combina con el hidrógeno del grupo carboxilo de un ácido graso, de esta manera el ácido graso se "ensambla" con el glicerol desprendiéndose agua (OH (del alcohol) + H (del carboxilo) ® H2O) . De la unión del glicerol con un ácido graso se forma un monoglicérido, con dos ácidos grasos tenemos un diglicérido, y con tres ácidos grasos tenemos un triglicérido.
Grasas y aceites:
Se diferencian uno del otro por que a temperatura ambiente los aceites son líquidos oleosos, esta característica está dada por que son triglicéridos no saturados, mientras que las grasas presentan ácidos grasos saturados. Ambos sirven de depósito de reserva de energía para células animales (grasas) y en vegetales (aceites). Estos compuestos son altamente energéticos, aproximadamente 9,3 kilocalorías por gramo. Cuando un organismo recibe energía asimilable en exceso, este puede almacenarla en forma de grasa, que podrá ser reutilizada posteriormente en la producción de energía, cuando el organismo lo necesite. En general, la grasa es almacenada en los adipocitos (células que forman el tejido adiposo) donde puede movilizarse para obtener energía cuando el ingreso calórico es menor que el gasto de calorías. Esta capa es utilizada en determinados animales como aislante térmico, como por ejemplo en mamíferos marino
Fosfolípidos:
Son los componentes primarios de las membranas celulares. En su estructura química podemos observar una molécula de glicerol, dos ácidos grasos, un grupo fosfato y un grupo de cabeza polar como por ejemplo la lecitina. Su fórmula general se representa de la siguiente manera:
Los fosfolípidos son anfipáticos, esto es que son simultaneamente hidrofílicos e hidrofóbicos. La "cabeza" de un fosfolípido es un grupo fosfato cargado negativamente y las dos "colas" son cadenas hidrocarbonadas fuertemente hidrofóbicas.
En las membranas celulares juegan un papel muy importante, ya que controlan la transferencia de sustancias hacia el interior o exterior de la célula. Una de las características de los fosfolípidos es que una parte de su estructura es soluble en agua (hidrofílica), mientras que la otra, es soluble en lípidos (hidrofóbica). La parte hidrofílica es en la que se encuentra el aminoalcohol o base nitrogenada. Esta característica estructural hace posible que los fosfolípidos participen en el intercambio de sustancias entre un sistema acuoso y un sistema lipídico, separando y aislando a los dos sistemas, a la vez que los mantiene juntos.
En medio acuoso las colas de los fosfolípidos tienden a disponerse en manera tal de formar un ambiente local hidrofóbico. Esto deja a los grupos fosfatos "de cara" al ambiente hidrofílico. Existen tres estructura que los fosfolípidos pueden formar en razón de naturaleza anfipática.
Esteroides:

Estructura de las Proteínas:











-es.svg/400px-Chloroplast_(borderless_version)-es.svg.png)






Es un grupo extenso de lípidos naturales o sintéticos con una diversidad de actividad fisiológica muy amplia. No se parecen a ningún otro lípido, se los ubica en esta clase por ser insolubles al agua. Todos los esteroides poseen cuatro anillos de carbono unido entre ellos, los que pueden presentar oxhidrilos o radicales. Entre los esteroides se encuentran:
El colesterol:
Existe en las membranas celulares (excepto las bacterianas y vegetales), un 25 % (peso en seco) de las membranas de los glóbulos rojos, y es un componente esencial de la vaina de mielina (cobertura de los axones de las neuronas). En cierta gente de edad avanzada forma depósitos grasos en el revestimiento interno de los vasos sanguíneos. Este depósitos pueden bloquear y reducir la elasticidad de los vasos, predisponiendo a la persona a sufrir: presión alta, ataques cardíacos, apoplejía.
Las hormonas sexuales y las de la corteza renal:
También son esteroides que se forman a partir del colesterol de los ovarios, testículos y otras glándulas.
Las prostaglandinas:
Son un grupo de sustancias químicas que poseen acciones hormonales y derivan de los ácidos grasos.
PROTEÍNAS:
Son las sustancias que componen las estructuras celulares y las herramientas que hacen posible las reacciones químicas del metabolismo celular. En la mayoría de los seres vivos (a excepción de las plantas que tienen más celulosa) representan más de un 50% de su peso en seco. Una bacteria puede tener cerca de 1000 proteínas diferentes, en una célula humana puede haber 10.000 clases de proteínas distintas.
Químicamente son macromoléculas, polímeros de aminoácidos (más de 100) dispuestos en una secuencia lineal, sin ramificaciones. Una secuencia de menos de 100 aminoácidos se denomina péptido.
Con la posibilidad de que 20 aminoácidos diferentes puedan ser ordenados en cualquier orden para conformar polipéptidos de cientos de aminoácidos, tienen el extraordinario potencial de producir una gran cantidad de variantes en su conformación. Esta variedad permite a las proteínas funciones tan refinadas como las de las enzimas que permiten el metabolismo celular. La bacteria Escherichia coli, uno de los organismos biológicos mas simples, tiene mas de 1000 proteínas diferentes trabajando a diferentes tiempos para catalizar las reacciones que sostienen a su vida.
Los Aminoácidos:
Los aminoácidos (aa) son moléculas orgánicas pequeñas con un grupo amino (NH2) y un grupo carboxilo (COOH). La gran cantidad de proteínas que se conocen están formadas únicamente por 20 aa diferentes. Se conocen otros 150 que no forman parte de las proteínas. Todos los aminoácidos tiene la misma formula general:
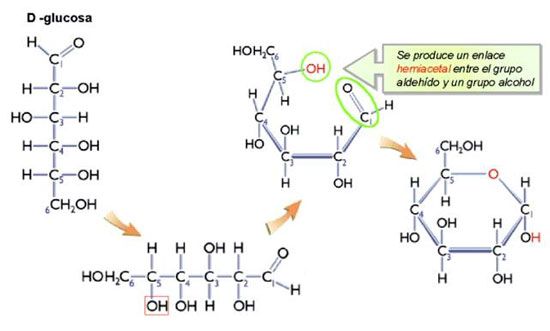
Generalmente, el número de AA que forman una proteína oscila entre 100 y 300. Los enlaces que participan en la estructura primaria de una proteína son covalentes: son los enlaces peptídicos. El enlace peptídico es un enlace amida que se forma entre el grupo carboxilo de una AA con el grupo amino de otro, con eliminación de una molécula de agua. Independientemente de la longitud de la cadena polipeptídica, siempre hay un extremo amino terminal y un extremo carboxilo terminal que permanecen intactos.
La unión peptídica:
Los aminoácidos se encuentran unidos linealmente por medio de uniones peptídicas. Estas uniones se forman por la reacción de síntesis (vía deshidratación) entre el grupo carboxilo del primer aminoácido con el grupo amino del segundo aminoácido.
Estructura de las Proteínas:
A primera vista podría pensarse en las proteínas como polímeros lineales de aminoácidos unidos entre sí por medio de enlaces peptídicos. Sin embargo, la secuencia lineal de aminoácidos puede adoptar múltiples conformaciones en el espacio.
Estructura primaria:
Las proteínas tiene múltiple niveles de estructura, la básica es la estructura primaria. La estructura primaria de una proteína es simplemente el orden de sus aminoácidos. Por convención el orden de escritura es siempre desde el grupo amino-terminal hasta el carboxilo final. Como consecuencia del establecimiento de enlaces peptídicos entre los distintos AA que forman la proteína se origina una cadena principal o "esqueleto" a partir del cual emergen las cadenas laterales de los aminoácidos.
Estructura secundaria:
La estructura secundaria de una proteína es la que adopta espacialmente. Existen ciertas estructuras repetitivas encontradas en las proteínas que permiten clasificarlas en dos tipos: hélice alfa y lámina beta.
Una hélice alfa es una apretada hélice formada por una cadena polipeptídica. La cadena polipetídica principal forma la estructura central, y las cadenas laterales se extienden por fuera de la hélice. El grupo carboxílo (CO) de un aminoácido n se une por puente hidrógeno al grupo amino (NH) de otro aminoácido que está tres residuos mas allá ( n + 4 ). De esta manera cada grupo CO y NH de la estructura central (columna vertebral o "backbone") se encuentra unido por puente hidrógeno.

B-Las láminas beta son el otro tipo de estructura secundaria. Pueden ser paralelas o antiparalelas. Las anti-paralelas generalmente se ven así: 

Estructura Terciaria:
A diferencia de la estructura secundaria, la estructura terciaria de la mayor parte de las proteínas es específica de cada molécula, además, determina su función. EL plegamiento terciario no es inmediato, primero se agrupan conjuntos de estructuras denominadas dominios que luego se articulan para formar la estructura terciaria definitiva. Este plegamiento está facilitado por uniones denominadas puentes disulfuro, -S-S- que se establecen entre los átomos de azufre del aminoácido cisteína.
Existen, sin embargo dos tipos de estructuras terciarias básicas:
- Proteínas fibrosas, insolubles en agua, como la alfa queratina o el colágeno, y
- Proteínas globulares, solubles en agua.
Estructura cuaternaria:
Solo está presente si hay mas de una cadena polipeptídica. Con varias cadenas polipeptídicas, la estructura cuaternaria representa su interconexión y organización. Esta es la imagen de la hemoglobina, una proteína con cuatro polipéptidos, dos alfa-globinas y dos beta globinas. En rojo se representa al grupo hem (complejo pegado a la proteína que contiene hierro, y sirve para transportar oxígeno).
Funciones de las proteínas:
Las proteínas determinan la forma y la estructura de las células y dirigen casi todos los procesos vitales. Las funciones de las proteínas son específicas de cada una de ellas y permiten a las células mantener su integridad, defenderse de agentes externos, reparar daños, controlar y regular funciones, etc. Todas las proteínas realizan su función de la misma manera: por unión selectiva a moléculas. Las proteínas estructurales se agregan a otras moléculas de la misma proteína para originar una estructura mayor. Sin embargo,otras proteínas se unen a moléculas distintas: los anticuerpos a los antígenos específicos, la hemoglobina al oxígeno, las enzimas a sus sustratos, los reguladores de la expresión génica al ADN, las hormonas a sus receptores específicos, etc.
A continuación se exponen algunos ejemplos de proteínas y las funciones que desempeñan:
Función estructural:
- Algunas proteínas constituyen estructuras celulares: ciertas glucoproteínas forman parte de las membranas celulares y actúan como receptores o facilitan el transporte de sustancias.
- Las histonas, forman parte de los cromosomas que regulan la expresión de los genes.
- Otras proteínas confieren elasticidad y resistencia a órganos y tejidos: el colágeno del tejido conjuntivo fibroso; la elastina del tejido conjuntivo elástico; la queratina de la epidermis.
- Las arañas y los gusanos de seda segregan fibroína para fabricar las telas de araña y los capullos de seda, respectivamente.
Función enzimática:
Las proteínas con función enzimática son las más numerosas y especializadas. Actúan como biocatalizadores de las reacciones químicas del metabolismo celular.
Función hormonal:
Algunas hormonas son de naturaleza proteica, como la insulina y el glucagón (que regulan los niveles de glucosa en sangre) o las hormonas segregadas por la hipófisis como la del crecimiento o la adrenocorticotrópica (que regula la síntesis de corticosteroides) o la calcitonina (que regula el metabolismo del calcio).
Función reguladora:
Algunas proteínas regulan la expresión de ciertos genes y otras regulan la división celular (como la ciclina).
Función homeostática:
Algunas mantienen el equilibrio osmótico y actúan junto con otros sistemas amortiguadores para mantener constante el pH del medio interno.
Función defensiva:
Las inmunoglogulinas actúan como anticuerpos frente a posibles antígenos. La trombina y el fibrinógeno contribuyen a la formación de coágulos sanguíneos para evitar hemorragias. Las mucinas tienen efecto germicida y protegen a las mucosas. Algunas toxinas bacterianas, como la del botulismo, o venenos de serpientes, son proteínas fabricadas con funciones defensivas.
Función de transporte:
- La hemoglobina transporta oxígeno en la sangre de los vertebrados.
- La hemocianina transporta oxígeno en la sangre de los invertebrados.
- La mioglobina transporta oxígeno en los músculos.
- Las lipoproteinas transportan lípidos por la sangre.
- Los citocromos transportan electrones.
Función contráctil:
- La actina y la miosina constituyen las miofibrillas responsables de la contracción muscular.
- La dineina está relacionada con el movimiento de cilios y flagelos.
Función de reserva:
La ovoalbúmina de la clara de huevo, la gliadina del grano de trigo y la hordeina de la cebada, constituyen la reserva de aminoácidos para el desarrollo del embrión.
La lactoalbúmina de la leche.
ÁCIDOS NUCLEÍCOS:
- Hay dos tipos de ácidos nucleicos (AN): el ácido desoxirribonucleico (ADN) y el ácido ribonucleico(ARN), y están presentes en todas las células. Su función biológica no quedó plenamente demostrada hasta que Avery y sus colaboradores demostraron en 1944 que el ADN era la molécula portadora de la información genética.
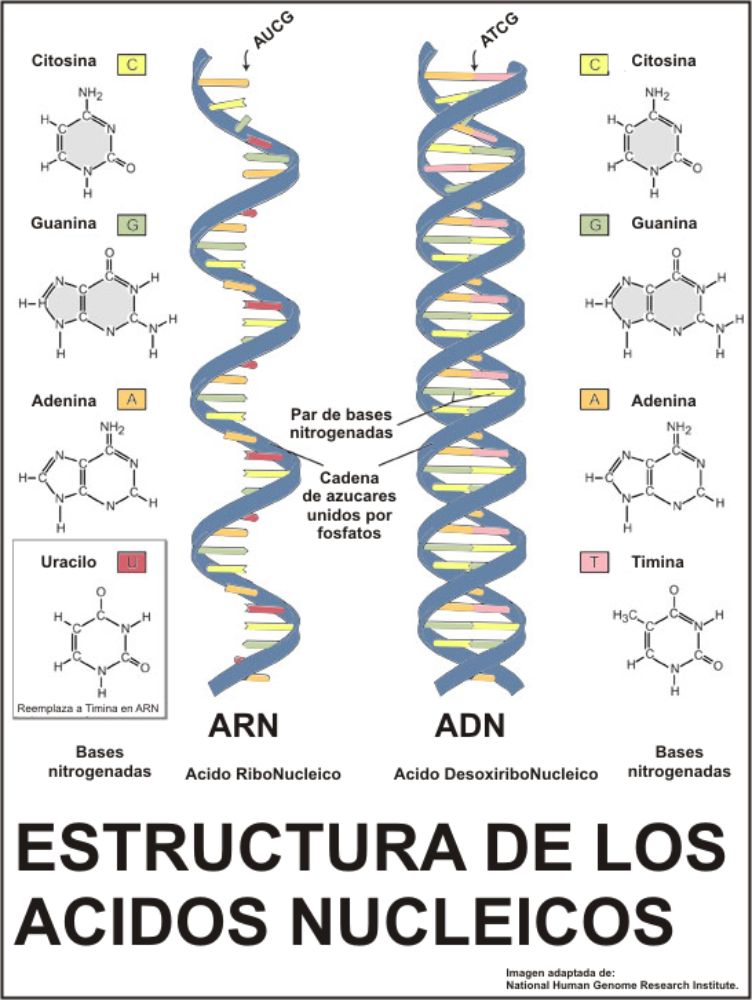
Las bases nitrogenadas pueden ser purinas: ADENINA y GUANINA. Las bases pirimidínicas son: CITOCINA, TIMINA y URACILO. La timina solo puede formar ADN y el uracilo solo está presente en el ARN.
Los nucleótidos se enlazan para formar los ácidos nucleicos o polinucleótidos a través de los carbones 3 y 5 de la molécula de azúcar tal como se muestra en la figura:
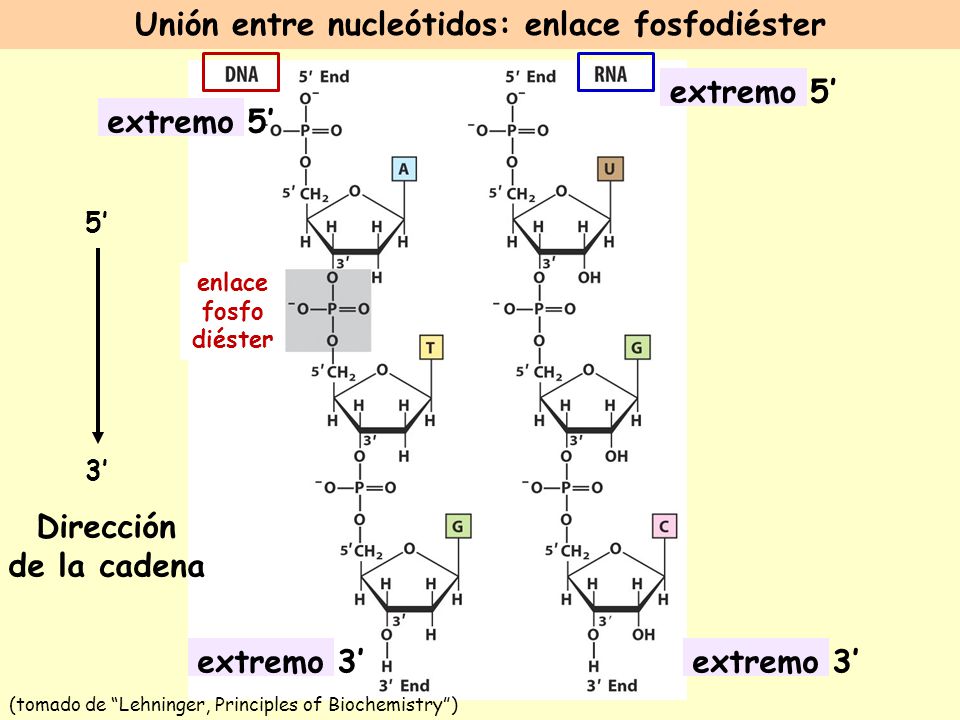
- Las bases nitrogenenadas que componen las moléculas del ADN son complementarias, así una A se enfrenta o complementa con una T, y una G se enfrenta o complementa con una C. A menudo los pares de bases son mencionados como A-T o G-C, adenina a timina y guanina a citosina. Raramente los libros o las personas usan los nombres completos de las bases. A-T están unidas por dos puentes Hidrógeno y C-G por tres.


Los nucleótidos se forman por la unión del C5' de la pentosa con el grupo fosfato formando un nucleótido monosfato. La cadena se va formando al enlazar los fosfatos al C3' de otro nucleótido. Así la cadena tiene un extremo 5´y un extremo 3´.
Estructura del ADN:
Algunos autores definen estructuras que denominan primarias, secundarias, etc. en orden de complejidad creciente, similar a las de las proteínas.
Las cuatro bases nitrogenadas del ADN se encuentran distribuidas a lo largo de la "columna vertebral" que conforman los azúcares con el ácido fosfórico en un orden particular, (la secuencia del ADN). La adenina (A) se empareja con la timina (T) mientras que la citosina (C) lo hace con la guanina.
La estructura primaria del ADN: está determinada por esta secuencia de bases ordenadas sobre la "columna" formada por los nucleósidos: azúcar + fosfato. Este orden es en realidad lo que se transmite de generación en generación (herencia).
Estructura secundaria: es el modelo postulado por Watson y Crick: la doble hélice, las dos hebras de ADN se mantienen unidas por los puentes hidrógenos entre las bases. Los pares de bases están formados siempre por una purina y una pirimidina, de forma que ambas cadenas están siempre equidistantes, a unos 11 Å una de la otra. Los pares de bases adoptan una disposición helicoidal en el núcleo central de la molécula, ya que presentan una rotación de 36º con respecto al par adyacente, de forma que hay 10 pares de bases por cada vuelta de la hélice. La A se empareja siempre con la T mediante dos puentes de hidrógeno, mientras que la C se empareja siempre con la G por medio de 3 puentes de hidrógeno.

En cada extremo de una doble hélice lineal de ADN, el extremo 3'-OH de una de las hebras es adyacente al extremo 5'-P (fosfato) de la otra. En otras palabras, las dos hebras son antiparalelas, es decir, tienen una orientación diferente. Por convención, la secuencia de bases de una hebra sencilla se escribe con el extremo 5'-P a la izquierda.
Estructura terciaria: es el súper arrollamiento que sufre la moléculas de ADN sobre proteínas globulares denominadas histonas con el fin de conformar los cromosomas:

En Procariotas (así como en las mitocondrias y cloroplastos eucariotas) el ADN se presenta como una doble cadena (de cerca de 1 mm de longitud), circular y cerrada, que toma el nombre de cromosoma bacteriano. Esta "gigantesca" molécula circular tiene un peso de 3 X 10 9d (daltons). No posee las histonas del cromosoma eucariota, pero se ha comprobado la existencia de proteínas y poliaminas de bajo peso molecular y de iones magnesio que cumplirían su función. El cromosoma bacteriano se encuentra altamente condensado y ordenado ("supercoiled" o superenrrollado). En virus, el ADN puede presentarse como una doble hélice cerrada, como una doble hélice abierta o simplemente como una única hebra lineal.
En los Eucariotas el ADN se encuentra localizado principalmente en el núcleo, apareciendo el superenrrollamiento (trenzamiento de la trenza) y la asociación con proteínas histónicas y no histónicas. El ADN se enrolla (dos vueltas) alrededor de un octeto de proteínas histónicas formando un nucleosoma, estos quedan separados por una secuencia de ADN de hasta 80 pares de bases, formando un "collar de perlas" o más correctamente denominado fibra de cromatina, siendo la estructura propia del núcleo interfásico, que no ha entrado en división. Este collar de nucleosomas vuelve a enrollarse y cada 6 nucleosomas constituyen un "paso de rosca" por medio de histoma H1 formando estructuras del tipo solenoide.
En el ciclo mitótico de las células eucariotas la cromatina se enrolla formando cromosomas, que son complejas asociaciones de ADN y proteínas .

Una célula típica contiene 10 veces más ARN que ADN. El azúcar presente en el ARN es la ribosa. Esto indica que en la posición 2' del anillo del azúcar hay un grupo hidroxilo (OH) libre. Por este motivo, el ARN es químicamente inestable, de forma que en una disolución acuosa se hidroliza fácilmente. En el ARN la base que se aparea con la A es U, a diferencia del ADN, en el cual la A se aparea con T.
Según las modernas teorías sobre el origen de la vida, parece bastante probable que el ARN fuese el primer biopolímero que apareció en la corteza terrestre durante el transcurso de la evolución.
Se distinguen varios tipos de RNA en función, sobre todo, de sus pesos moleculares:
- RNA mensajero (RNAm): se sintetiza sobre un molde de ADN por el proceso de transcripción por el cual se copia el ARN a partir del molde del ADN, pasa al citoplasma y sirve de pauta para la síntesis de proteínas (traducción).
RNA robosómico (RNAr): el RNA ribosómico (RNAr) está presente en los ribosomas, orgánulos intracelulares implicados en la síntesis de proteínas. Su función es leer los RNAm y formar la proteína correspondiente.
RNA de transferencia: son cadenas cortas de una estructura básica, que pueden unirse específicamente a determinados aminoácidos.
Se invita al lector a observar el siguiente vídeo de la temática tratada, para tener mayor conocimiento sobre los biocompuestos.
También se podrá observar los temas tratados en las siguientes diapositivas:
ACTIVIDAD:
1. TALLER SOBRE BIOCOMPUESTOS
La presente actividad tiene como objeto desarrollar aspectos educativos y conceptuales a cerca de las moléculas que componen los sistemas vivos (biomoléculas). En este caso particular, se abordarán conceptos relacionados con los glúcidos, lípidos, proteínas y ácidos nucleicos: su estructura, importancia funcional y los aspectos especiales de estas moléculas que las hace importante en la naturaleza.
Para el caso, individualmente, cada estudiante deberá:
1. Desarrollar el cuestionario propuesto.
3. El trabajo deberá ser elaborado con los siguientes elementos formales:
· Portada
· Introducción
· Desarrollo
· Conclusiones
· Bibliografía (haga uso de bibliografía especializada)
4. El trabajo se entregará en medio magnético, formato PDF, letra arial o times new roman número 12, interlineado fuera de cuadro 1,5.
CUESTIONARIO
1. Elabore una tabla comparativa que relacione de manera general la composición y la función biológica que poseen los glúcidos, los lípidos, las proteínas y los ácidos nucleicos.
2. Realice una tabla en la cual relacione la clasificación de los glúcidos de acuerdo a su complejidad.
3. Explique por qué los lípidos son moléculas anfipáticas. ¿Qué importancia tiene esto para las células?
4. Usando gráficas, represente el enlace peptídico que ocurre entre los aminoácidos para formar un dipéptido.
5. Describa de manera breve los diferentes tipos de estructuras de las proteínas, represéntelas con imágenes.
6. En media cuartilla argumente las diferencias que existen entre una proteína fibrosa como la colágena y una proteína globular como la hemoglobina.
7. Explique de manera clara y resumida, en qué consiste la estructura secundaria del ADN. Represéntela gráficamente.
CLASE 2: CARACTERÍSTICAS ESTRUCTURALES Y FUNCIONALES DE LAS CÉLULAS
1. Generalidades.
La célula (del latín cellula, diminutivo de cella, ‘celda’) es la unidad morfológica y funcional de todo ser vivo. De hecho, la célula es el elemento de menor tamaño que puede considerarse vivo. De este modo, puede clasificarse a los organismos vivos según el número de células que posean: si solo tienen una, se les denomina unicelulares (como pueden ser los protozoos o las bacterias, organismos microscópicos); si poseen más, se les llama pluricelulares. En estos últimos el número de células es variable: de unos pocos cientos, como en algunos nematodos, a cientos de billones como en el caso del ser humano. Las células suelen poseer un tamaño de 10 µm y una masa de 1 ng (nano gramo), si bien existen células mucho mayores.
La teoría celular, propuesta en 1838 para los vegetales y en 1839 para los animales, por Matthias Jakob Schleiden y Theodor Schwann, postula que todos los organismos están compuestos por células, y que todas las células derivan de otras precedentes. De este modo, todas las funciones vitales emanan de la maquinaria celular y de la interacción entre células adyacentes; además, la tenencia de la información genética, base de la herencia, en su ADN permite la transmisión de aquella de generación en generación.
La aparición del primer organismo vivo sobre la Tierra suele asociarse al nacimiento de la primera célula. Si bien existen muchas hipótesis que especulan cómo ocurrió, usualmente se describe que el proceso se inició gracias a la transformación de moléculas inorgánicas en orgánicas bajo unas condiciones ambientales adecuadas; tras esto, dichas biomoléculas se asociaron dando lugar a entes complejos capaces de autorreplicarse. Existen posibles evidencias fósiles de estructuras celulares en rocas datadas en torno a 4 o 3,5 miles de millones de años (giga-años o Ga.). Se han encontrado evidencias muy fuertes de formas de vida unicelulares fosilizadas en microestructuras en rocas de la formación Strelley Pool, en Australia Occidental, con una antigüedad de 3,4 Ga. Se trataría de los fósiles de células más antiguos encontrados hasta la fecha. Evidencias adicionales muestran que su metabolismo sería anaerobio y basado en el sulfuro.
2. Características.
Las células, como sistemas termodinámicos complejos, poseen una serie de elementos estructurales y funcionales comunes que posibilitan su supervivencia; no obstante, los distintos tipos celulares presentan modificaciones de estas características comunes que permiten su especialización funcional y, por ello, la ganancia de complejidad. De este modo, las células permanecen altamente organizadas a costa de incrementar la entropía del entorno, uno de los requisitos de la vida.
2.1. Características estructurales:
- Individualidad: Todas las células están rodeadas de una envoltura (que puede ser una bicapa lipídica desnuda, en células animales; una pared de polisacárido, en hongos y vegetales; una membrana externa y otros elementos que definen una pared compleja, en bacterias Gram negativas; una pared de peptidoglicano, en bacterias Gram positivas; o una pared de variada composición, en arqueas) que las separa y comunica con el exterior, que controla los movimientos celulares y que mantiene el potencial de membrana.
- Contienen un medio interno acuoso: el citosol, que forma la mayor parte del volumen celular y en el que están inmersos los orgánulos celulares.
- Poseen material genético: en forma de ADN, el material hereditario de los genes, que contiene las instrucciones para el funcionamiento celular, así como ARN, a fin de que el primero se exprese.
- Tienen enzimas y otras proteínas: que sustentan, junto con otras biomoléculas, un metabolismo activo.
2.2. Características funcionales:
Las células vivas son un sistema bioquímico complejo. Las características que permiten diferenciar las células de los sistemas químicos no vivos son:
- Nutrición: las células toman sustancias del medio, las transforman de una forma a otra, liberan energía y eliminan productos de desecho, mediante el metabolismo.
- Crecimiento y multiplicación: las células son capaces de dirigir su propia síntesis. A consecuencia de los procesos nutricionales, una célula crece y se divide, formando dos células, en una célula idéntica a la célula original, mediante la división celular.
- Diferenciación: muchas células pueden sufrir cambios de forma o función en un proceso llamado diferenciación celular. Cuando una célula se diferencia, se forman algunas sustancias o estructuras que no estaban previamente formadas y otras que lo estaban dejan de formarse. La diferenciación es a menudo parte del ciclo celular en que las células forman estructuras especializadas relacionadas con la reproducción, la dispersión o la supervivencia.
- Señalización: las células responden a estímulos químicos y físicos tanto del medio externo como de su interior y, en el caso de células móviles, hacia determinados estímulos ambientales o en dirección opuesta mediante un proceso que se denomina quimiotaxis. Además, frecuentemente las células pueden interaccionar o comunicar con otras células, generalmente por medio de señales o mensajeros químicos, como hormonas, neurotransmisores, factores de crecimiento...
- Evolución: a diferencia de las estructuras inanimadas, los organismos unicelulares y pluricelulares evolucionan. Esto significa que hay cambios hereditarios (que ocurren a baja frecuencia en todas las células de modo regular) que pueden influir en la adaptación global de la célula o del organismo superior de modo positivo o negativo. El resultado de la evolución es la selección de aquellos organismos mejor adaptados a vivir en un medio particular.
Las propiedades celulares no tienen por qué ser constantes a lo largo del desarrollo de un organismo: evidentemente, el patrón de expresión de los genes varía en respuesta a estímulos externos, además de factores endógenos. Un aspecto importante a controlar es la pluripotencialidad, característica de algunas células que les permite dirigir su desarrollo hacia un abanico de posibles tipos celulares. En metazoos, la genética subyacente a la determinación del destino de una célula consiste en la expresión de determinados factores de transcripción específicos del linaje celular al cual va a pertenecer, así como a modificaciones epigenéticas. Además, la introducción de otro tipo de factores de transcripción mediante ingeniería genética en células somáticas basta para inducir la mencionada pluripotencialidad, luego este es uno de sus fundamentos moleculares.
3. Tamaño, forma y función.
El tamaño y la forma de las células depende de sus elementos más periféricos (por ejemplo, la pared, si la hubiere) y de su andamiaje interno (es decir, el citoesqueleto). Además, la competencia por el espacio tisular provoca una morfología característica: por ejemplo, las células vegetales, poliédricas in vivo, tienden a ser esféricas in vitro. Incluso pueden existir parámetros químicos sencillos, como los gradientes de concentración de una sal, que determinen la aparición de una forma compleja.
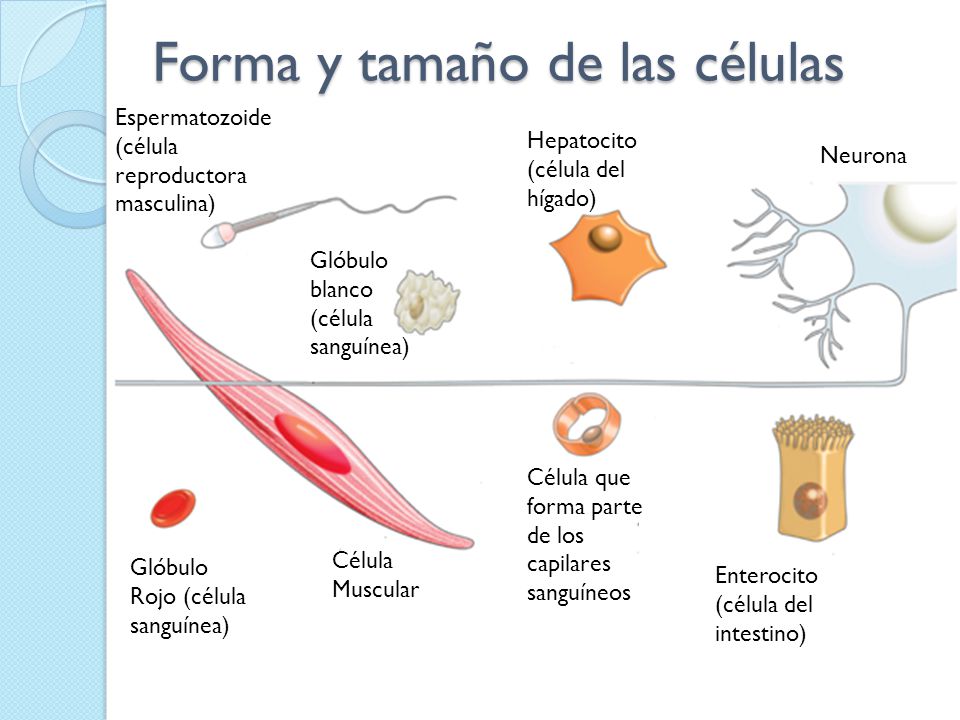
En cuanto al tamaño, la mayoría de las células son microscópicas, es decir, no son observables a simple vista. (un milímetro cúbico de sangre puede contener unos cinco millones de células), a pesar de ser muy pequeñas el tamaño de las células es extremadamente variable. La célula más pequeña observada, en condiciones normales, corresponde a Mycoplasma genitalium, de 0,2 μm, encontrándose cerca del límite teórico de 0,17 μm. Existen bacterias con 1 y 2 μm de longitud. Las células humanas son muy variables: hematíes de 7 micras, hepatocitos con 20 micras, espermatozoides de 53 μm, óvulos de 150 μm e, incluso, algunas neuronas de en torno a un metro. En las células vegetales los granos de polen pueden llegar a medir de 200 a 300 μm.
Respecto a las células de mayor tamaño; por ejemplo los xenofióforos, son foraminíferos unicelulares que han desarrollado un gran tamaño, los cuales alcanzar tamaños macroscópicos (Syringammina fragilissima alcanza los 20 cm de diámetro).
Para la viabilidad de la célula y su correcto funcionamiento siempre se debe tener en cuenta la relación superficie-volumen. Puede aumentar considerablemente el volumen de la célula y no así su superficie de intercambio de membrana lo que dificultaría el nivel y regulación de los intercambios de sustancias vitales para la célula.
Respecto de su forma, las células presentan una gran variabilidad, e, incluso, algunas no la poseen bien definida o permanente. Pueden ser: fusiformes (forma de huso), estrelladas, prismáticas, aplanadas, elípticas, globosas o redondeadas, etc. Algunas tienen una pared rígida y otras no, lo que les permite deformar la membrana y emitir prolongaciones citoplasmáticas (pseudópodos) para desplazarse o conseguir alimento. Hay células libres que no muestran esas estructuras de desplazamiento pero poseen cilios o flagelos, que son estructuras derivadas de un orgánulo celular (el centrosoma) que dota a estas células de movimiento. De este modo, existen multitud de tipos celulares, relacionados con la función que desempeñan; por ejemplo:
- Células contráctiles que suelen ser alargadas, como las fibras musculares.
- Células con finas prolongaciones, como las neuronas que transmiten el impulso nervioso.
- Células con microvellosidades o con pliegues, como las del intestino para ampliar la superficie de contacto y de intercambio de sustancias.
- Células cúbicas, prismáticas o aplanadas como las epiteliales que recubren superficies como las losas de un pavimento.
4. La célula procariótica.
Las células procariotas son pequeñas y menos complejas que las eucariotas. Contienen ribosomas pero carecen de sistemas de endomembranas (esto es, orgánulos delimitados por membranas biológicas, como puede ser el núcleo celular). Por ello poseen el material genético en el citosol. Sin embargo, existen excepciones: algunas bacterias fotosintéticas poseen sistemas de membranas internos. También en el Filo Planctomycetes existen organismos como Pirellula que rodean su material genético mediante una membrana intracitoplasmática y Gemmata obscuriglobus que lo rodea con doble membrana. Esta última posee además otros compartimentos internos de membrana, posiblemente conectados con la membrana externa del nucleoide y con la membrana plasmática, que no está asociada a peptidoglucano. Estudios realizados en 2017, demuestran otra particularidad de Gemmata: presenta estructuras similares al poro nuclear, en la membrana que rodea su cuerpo nuclear.
Por lo general podría decirse que los procariotas carecen de citoesqueleto. Sin embargo se ha observado que algunas bacterias, como Bacillus subtilis, poseen proteínas tales como MreB y mbl que actúan de un modo similar a la actina y son importantes en la morfología celular.
De gran diversidad, los procariotas sustentan un metabolismo extraordinariamente complejo, en algunos casos exclusivo de ciertos taxa, como algunos grupos de bacterias, lo que incide en su versatilidad ecológica. Los procariotas se clasifican, según Carl Woese, en arqueas y bacterias.
4.1 Arqueas:
Las arqueas, al igual que las bacterias, no tienen membranas internas que delimiten orgánulos. Como todos los organismos presentan ribosomas, pero a diferencia de los encontrados en las bacterias que son sensibles a ciertos agentes antimicrobianos, los de las arqueas, más cercanos a los eucariotas, no lo son. La membrana celular tiene una estructura similar a la de las demás células, pero su composición química es única, con enlaces tipo éter en sus lípidos. Casi todas las arqueas poseen una pared celular (algunos Thermoplasma son la excepción) de composición característica, por ejemplo, no contienen peptidoglicano (mureína), propio de bacterias. No obstante pueden clasificarse bajo la tinción de Gram, de vital importancia en la taxonomía de bacterias; sin embargo, en arqueas, poseedoras de una estructura de pared en absoluto común a la bacteriana, dicha tinción es aplicable pero carece de valor taxonómico. El orden Methanobacteriales tiene una capa de pseudomureína, que provoca que dichas arqueas respondan como positivas a la tinción de Gram.
Como en casi todos los procariotas, las células de las arqueas carecen de núcleo, y presentan un solo cromosoma circular. Existen elementos extracromosómicos, tales como plásmidos. Sus genomas son de pequeño tamaño, sobre 2-4 millones de pares de bases. También es característica la presencia de ARN polimerasas de constitución compleja y un gran número de nucleótidos modificados en los ácidos ribonucleicos ribosomales. Por otra parte, su ADN se empaqueta en forma de nucleosomas, como en los eucariotas, gracias a proteínas semejantes a las histonas y algunos genes poseen intrones. Pueden reproducirse por fisión binaria o múltiple, fragmentación o gemación.
4.2 Bacterias:
Las bacterias son organismos relativamente sencillos, de dimensiones muy reducidas, de apenas unas micras en la mayoría de los casos. Como otros procariotas, carecen de un núcleo delimitado por una membrana, aunque presentan un nucleoide, una estructura elemental que contiene una gran molécula generalmente circular de ADN. Carecen de núcleo celular y demás orgánulos delimitados por membranas biológicas. En el citoplasma se pueden apreciar plásmidos, pequeñas moléculas circulares de ADN que coexisten con el nucleoide y que contienen genes: son comúnmente usados por las bacterias en la parasexualidad (reproducción sexual bacteriana). El citoplasma también contiene ribosomas y diversos tipos de gránulos. En algunos casos, puede haber estructuras compuestas por membranas, generalmente relacionadas con la fotosíntesis.
Poseen una membrana celular compuesta de lípidos, en forma de una bicapa y sobre ella se encuentra una cubierta en la que existe un polisacárido complejo denominado peptidoglicano; dependiendo de su estructura y subsecuente su respuesta a la tinción de Gram, se clasifica a las bacterias en Gram positivas y Gram negativas. El espacio comprendido entre la membrana celular y la pared celular (o la membrana externa, si esta existe) se denomina espacio periplásmico. Algunas bacterias presentan una cápsula. Otras son capaces de generar endosporas (estadios latentes capaces de resistir condiciones extremas) en algún momento de su ciclo vital. Entre las formaciones exteriores propias de la célula bacteriana destacan los flagelos (de estructura completamente distinta a la de los flagelos eucariotas) y los pili (estructuras de adherencia y relacionadas con la parasexualidad).
La mayoría de las bacterias disponen de un único cromosoma circular y suelen poseer elementos genéticos adicionales, como distintos tipos de plásmidos. Su reproducción, binaria y muy eficiente en el tiempo, permite la rápida expansión de sus poblaciones, generándose un gran número de células que son virtualmente clones, esto es, idénticas entre sí.

5. La célula eucariótica.
Las células eucariotas son el exponente de la complejidad celular actual. Presentan una estructura básica relativamente estable caracterizada por la presencia de distintos tipos de orgánulos intracitoplasmáticos especializados, entre los cuales destaca el núcleo, que alberga el material genético. Especialmente en los organismos pluricelulares, las células pueden alcanzar un alto grado de especialización. Dicha especialización o diferenciación es tal que, en algunos casos, compromete la propia viabilidad del tipo celular en aislamiento. Así, por ejemplo, las neuronas dependen para su supervivencia de las células gliales.
Por otro lado, la estructura de la célula varía dependiendo de la situación taxonómica del ser vivo: de este modo, las células vegetales difieren de las animales, así como de las de los hongos. Por ejemplo, las células animales carecen de pared celular, son muy variables, no tiene plastos, puede tener vacuolas pero no son muy grandes y presentan centríolos (que son agregados de microtúbulos cilíndricos que contribuyen a la formación de los cilios y los flagelos y facilitan la división celular). Las células de los vegetales, por su lado, presentan una pared celular compuesta principalmente de celulosa, disponen de plastos como cloroplastos (orgánulo capaz de realizar la fotosíntesis), cromoplastos (orgánulos que acumulan pigmentos) o leucoplastos (orgánulos que acumulan el almidón fabricado en la fotosíntesis), poseen vacuolas de gran tamaño que acumulan sustancias de reserva o de desecho producidas por la célula y finalmente cuentan también con plasmodesmos, que son conexiones citoplasmáticas que permiten la circulación directa de las sustancias del citoplasma de una célula a otra, con continuidad de sus membranas plasmáticas.


5.1 Compartimentos:
Las células son entes dinámicos, con un metabolismo celular interno de gran actividad cuya estructura es un flujo entre rutas anastomosadas. Un fenómeno observado en todos los tipos celulares es la compartimentalización, que consiste en una heterogeneidad que da lugar a entornos más o menos definidos (rodeados o no mediante membranas biológicas) en las cuales existe un microentorno que aglutina a los elementos implicados en una ruta biológica. Esta compartimentalización alcanza su máximo exponente en las células eucariotas, las cuales están formadas por diferentes estructuras y orgánulos que desarrollan funciones específicas, lo que supone un método de especialización espacial y temporal. No obstante, células más sencillas, como los procariotas, ya poseen especializaciones semejantes.
5.2 Membrana plasmática y superficie celular:
La composición de la membrana plasmática varía entre células dependiendo de la función o del tejido en la que se encuentre, pero posee elementos comunes. Está compuesta por una doble capa de fosfolípidos, por proteínas unidas no covalentemente a esa bicapa, y por glúcidos unidos covalentemente a lípidos o proteínas. Generalmente, las moléculas más numerosas son las de lípidos; sin embargo, las proteínas, debido a su mayor masa molecular, representan aproximadamente el 50 % de la masa de la membrana.
Un modelo que explica el funcionamiento de la membrana plasmática es el modelo del mosaico fluido, de J. S. Singer y Garth Nicolson (1972), que desarrolla un concepto de unidad termodinámica basada en las interacciones hidrófobas entre moléculas y otro tipo de enlaces no covalentes.
Esquema de una membrana celular. Se observa la bicapa de fosfolípidos, las proteínas y otras moléculas asociadas que permiten las funciones inherentes a este orgánulo.
Dicha estructura de membrana sustenta un complejo mecanismo de transporte, que posibilita un fluido intercambio de masa y energía entre el entorno intracelular y el externo. Además, la posibilidad de transporte e interacción entre moléculas de células aledañas o de una célula con su entorno faculta a estas poder comunicarse químicamente, esto es, permite la señalización celular. Neurotransmisores, hormonas, mediadores químicos locales afectan a células concretas modificando el patrón de expresión génica mediante mecanismos de transducción de señal.
Sobre la bicapa lipídica, independientemente de la presencia o no de una pared celular, existe una matriz que puede variar, de poco conspicua, como en los epitelios, a muy extensa, como en el tejido conjuntivo. Dicha matriz, denominada glucocalix (glicocáliz), rica en líquido tisular, glucoproteínas, proteoglicanos y fibras, también interviene en la generación de estructuras y funciones emergentes, derivadas de las interacciones célula-célula.
5.3 Estructura y expresión génica:
Las células eucariotas poseen su material genético en, generalmente, un solo núcleodelimitado por una envoltura consistente en dos bicapas lipídicas atravesadas por numerosos poros nucleares y en continuidad con el retículo endoplasmático. En su interior, se encuentra el material genético, el ADN, observable, en las células en interfase, como cromatina de distribución heterogénea. A esta cromatina se encuentran asociadas multitud de proteínas, entre las cuales destacan las histonas, así como ARN, otro ácido nucleico.
Dicho material genético se encuentra inmerso en una actividad continua de regulación de la expresión génica; las ARN polimerasastranscriben ARN mensajero continuamente, que, exportado al citosol, es traducido a proteína, de acuerdo a las necesidades fisiológicas. Asimismo, dependiendo del momento del ciclo celular, dicho ADN puede entrar en replicación, como paso previo a la mitosis. No obstante, las células eucarióticas poseen material genético extranuclear: concretamente, en mitocondrias y plastos, si los hubiere; estos orgánulos conservan una independencia genética parcial del genoma nuclear.
Dicho material genético se encuentra inmerso en una actividad continua de regulación de la expresión génica; las ARN polimerasastranscriben ARN mensajero continuamente, que, exportado al citosol, es traducido a proteína, de acuerdo a las necesidades fisiológicas. Asimismo, dependiendo del momento del ciclo celular, dicho ADN puede entrar en replicación, como paso previo a la mitosis. No obstante, las células eucarióticas poseen material genético extranuclear: concretamente, en mitocondrias y plastos, si los hubiere; estos orgánulos conservan una independencia genética parcial del genoma nuclear.
5.4 Síntesis y degradación de macromoléculas:
Dentro del citosol, esto es, la matriz acuosa que alberga a los orgánulos y demás estructuras celulares, se encuentran inmersos multitud de tipos de maquinaria de metabolismo celular: orgánulos, inclusiones, elementos del citoesqueleto, enzimas. De hecho, estas últimas corresponden al 20 % de las enzimas totales de la célula.
- Ribosoma: Los ribosomas, visibles al microscopio electrónico como partículas esféricas, son complejos supramoleculares encargados de ensamblar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero. Elaborados en el núcleo, desempeñan su función de síntesis de proteínas en el citoplasma. Están formados por ARN ribosómico y por diversos tipos de proteínas. Estructuralmente, tienen dos subunidades. En las células, estos orgánulos aparecen en diferentes estados de disociación. Cuando están completos, pueden estar aislados o formando grupos (polisomas). También pueden aparecer asociados al retículo endoplasmático rugoso o a la envoltura nuclear.

Ribosomas: subunidad mayor en rojo, sub unidad menor en azul
- Retículo endoplasmático: El retículo endoplasmático es orgánulo vesicular interconectado que forma cisternas, tubos aplanados y sáculos comunicados entre sí. Intervienen en funciones relacionadas con la síntesis proteica, glicosilación de proteínas, metabolismo de lípidos y algunos esteroides, detoxificación, así como el tráfico de vesículas. En células especializadas, como las miofibrillas o células musculares, se diferencia en el retículo sarcoplásmico, orgánulo decisivo para que se produzca la contracción muscular.
- Aparato de Golgi: El aparato de Golgi es un orgánulo formado por apilamientos de sáculos denominados dictiosomas, si bien, como ente dinámico, estos pueden interpretarse como estructuras puntuales fruto de la coalescencia de vesículas. Recibe las vesículas del retículo endoplasmático rugoso que han de seguir siendo procesadas. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos y la síntesis de polisacáridos de la matriz extracelular. Posee tres compartimientos; uno proximal al retículo endoplasmático, denominado «compartimento cis», donde se produce la fosforilación de las manosas de las enzimas que han de dirigirse al lisosoma; el «compartimento intermedio», con abundantes manosidasas y N-acetil-glucosamina transferasas; y el «compartimento o red trans», el más distal, donde se transfieren residuos de galactosa y ácido siálico, y del que emergen las vesículas con los diversos destinos celulares.
- Lisosoma: Los lisosomas son orgánulos que albergan multitud de enzimas hidrolíticas. De morfología muy variable, no se ha demostrado su existencia en células vegetales. Una característica que agrupa a todos los lisosomas es la posesión de hidrolasas ácidas: proteasas, nucleasas, glucosidasas, lisozima, arilsulfatasas, lipasas, fosfolipasas y fosfatasas. Procede de la fusión de vesículas procedentes del aparato de Golgi, que, a su vez, se fusionan en un tipo de orgánulo denominado endosoma temprano, el cual, al acidificarse y ganar en enzimas hidrolíticos, pasa a convertirse en el lisosoma funcional. Sus funciones abarcan desde la degradación de macromoléculas endógenas o procedentes de la fagocitosis a la intervención en procesos de apoptosis.
- La vacuola regula el estado de turgencia de la célula vegetal.
- Vacuola vegetal: Las vacuolas vegetales, numerosas y pequeñas en células meristemáticas y escasas y grandes en células diferenciadas, son orgánulos exclusivos de los representantes del mundo vegetal. Inmersas en el citosol, están delimitadas por el tonoplasto, una membrana lipídica. Sus funciones son: facilitar el intercambio con el medio externo, mantener la turgencia celular, la digestión celular y la acumulación de sustancias de reserva y subproductos del metabolismo.
Vacuolas de células epidérmicas de tradescantia: Se observa que contienen pigmentos de antocianinas en su interior y se han retraído al centro de la célula al sufrir un proceso de plasmólisis.
- Inclusión citoplasmática: Las inclusiones son acúmulos nunca delimitados por membrana de sustancias de diversa índole, tanto en células vegetales como animales. Típicamente se trata de sustancias de reserva que se conservan como acervo metabólico: almidón, glucógeno, triglicéridos, proteínas... aunque también existen de pigmentos.
5.5 Conversión energética:
El metabolismo celular está basado en la transformación de unas sustancias químicas denominadas metabolitos. Dichas reacciones químicas transcurren catalizadas mediante enzimas. Si bien buena parte del metabolismo sucede en el citosol, como la glucólisis, existen procesos específicos realizados en orgánulos.
- Mitocondria: las mitocondrias son orgánulos de aspecto, número y tamaño variable que intervienen en el ciclo de Krebs, fosforilación oxidativa y en la cadena de transporte de electrones de la respiración. Presentan una doble membrana, externa e interna, que dejan entre ellas un espacio perimitocondrial; la membrana interna, plegada en crestas hacia el interior de la matriz mitocondrial, posee una gran superficie. En su interior posee generalmente una sola molécula de ADN, el genoma mitocondrial, típicamente circular, así como ribosomas más semejantes a los bacterianos que a los eucariotas. Según la teoría endosimbiótica, se asume que la primera protomitocondria era un tipo de proteobacteria.
- Cloroplasto: los cloroplastos son los orgánulos celulares que en los organismos eucariotas fotosintéticos se ocupan de la fotosíntesis. Están limitados por una envoltura formada por dos membranas concéntricas y contienen vesículas, los tilacoides, donde se encuentran organizados los pigmentos y demás moléculas implicadas en la conversión de la energía lumínica en energía química. Además de esta función, los plastidios intervienen en el metabolismo intermedio, produciendo energía y poder reductor, sintetizando bases púricas y pirimidínicas, algunos aminoácidos y todos los ácidos grasos. Además, en su interior es común la acumulación de sustancias de reserva, como el almidón. Se considera que poseen analogía con las cianobacterias.
- Peroxisoma: Los peroxisomas son orgánulos muy comunes en forma de vesículas que contienen abundantes enzimas de tipo oxidasa y catalasa; de tan abundantes, es común que cristalicen en su interior. Estas enzimas cumplen funciones de detoxificación celular. Otras funciones de los peroxisomas son: las oxidaciones flavínicas generales, el catabolismo de las purinas, la beta-oxidación de los ácidos grasos, el ciclo del glioxilato, el metabolismo del ácido glicólico y la detoxificación en general. Se forman de vesículas procedentes del retículo endoplasmático.
5.6 Citoesqueleto:
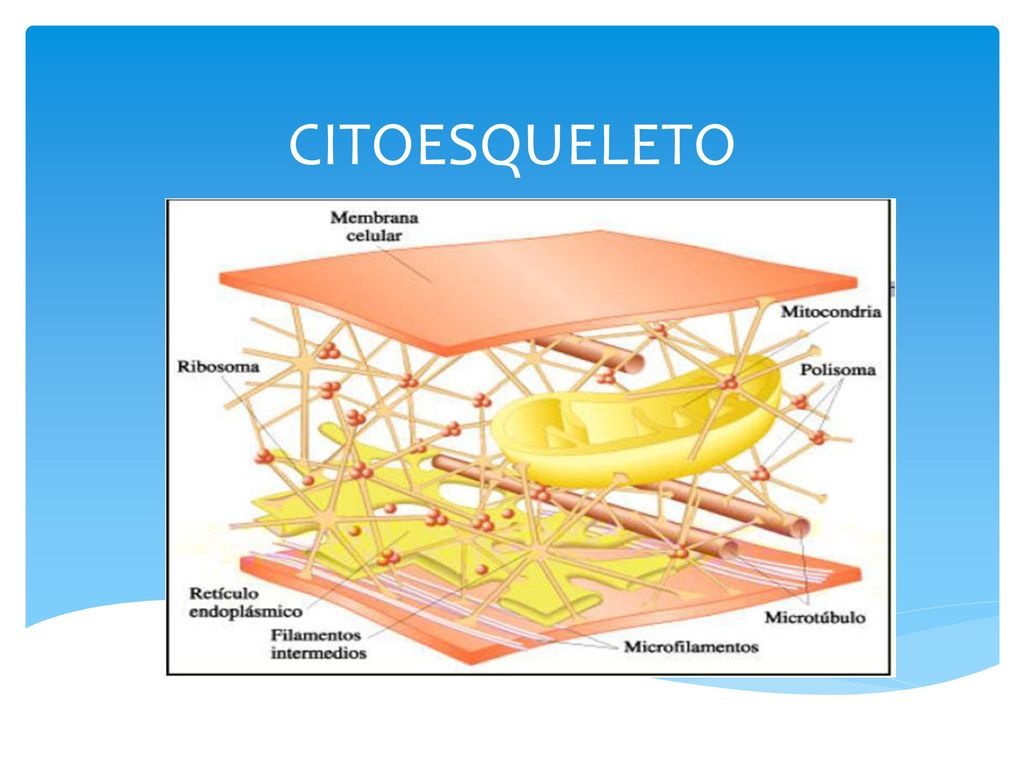
Las células poseen un andamiaje que permite el mantenimiento de su forma y estructura, pero más aún, este es un sistema dinámico que interactúa con el resto de componentes celulares generando un alto grado de orden interno. Dicho andamiaje está formado por una serie de proteínas que se agrupan dando lugar a estructuras filamentosas que, mediante otras proteínas, interactúan entre ellas dando lugar a una especie de retículo. El mencionado andamiaje recibe el nombre de citoesqueleto, y sus elementos mayoritarios son: los microtúbulos, los microfilamentos y los filamentos intermedios.
- Microfilamentos: Los microfilamentos o filamentos de actina están formados por una proteína globular, la actina, que puede polimerizar dando lugar a estructuras filiformes. Dicha actina se expresa en todas las células del cuerpo y especialmente en las musculares ya que está implicada en la contracción muscular, por interacción con la miosina. Además, posee lugares de unión a ATP, lo que dota a sus filamentos de polaridad. Puede encontrarse en forma libre o polimerizarse en microfilamentos, que son esenciales para funciones celulares tan importantes como la movilidad y la contracción de la célula durante la división celular.
- Microtúbulos: Los microtúbulos son estructuras tubulares de 25 nm de diámetro exterior y unos 12 nm de diámetro interior, con longitudes que varían entre unos pocos nanómetros a micrómetros, que se originan en los centros organizadores de microtúbulos y que se extienden a lo largo de todo el citoplasma. Se hallan en las células eucariotas y están formadas por la polimerización de un dímero de dos proteínas globulares, la alfa y la beta tubulina. Las tubulinas poseen capacidad de unir GTP (Guanosin-trifosfato). Los microtúbulos intervienen en diversos procesos celulares que involucran desplazamiento de vesículas de secreción, movimiento de orgánulos, transporte intracelular de sustancias, así como en la división celular (mitosis y meiosis) y que, junto con los microfilamentos y los filamentos intermedios, forman el citoesqueleto. Además, constituyen la estructura interna de los cilios y los flagelos.
- Filamentos intermedios: Los filamentos intermedios son componentes del citoesqueleto. Formados por agrupaciones de proteínas fibrosas, su nombre deriva de su diámetro, de 10 nm, menor que el de los microtúbulos, de 24 nm, pero mayor que el de los microfilamentos, de 7 nm. Son ubicuos en las células animales, y no existen en plantas ni hongos. Forman un grupo heterogéneo, clasificado en cinco familias: las queratinas, en células epiteliales; los neurofilamentos, en neuronas; los gliofilamentos, en células gliales; la desmina, en músculo liso y estriado; y la vimentina, en células derivadas del mesénquima.
- Centríolos: Los centríolos son una pareja de estructuras que forman parte del citoesqueleto de células animales. Semejantes a cilindros huecos, están rodeados de un material proteico denso llamado material pericentriolar; todos ellos forman el centrosoma o centro organizador de microtúbulos que permiten la polimerización de microtúbulos de dímeros de tubulina que forman parte del citoesqueleto. Los centríolos se posicionan perpendicularmente entre sí. Sus funciones son participar en la mitosis, durante la cual generan el huso acromático, y en la citocinesis, así como, se postula, intervenir en la nucleación de microtúbulos.
Se invita a observar el siguiente vídeo para contextualizar el tema tratado.
ACTIVIDAD
Los equipos de trabajo realizarán una presentación en PREZI sobre la célula eucariótica. En dicha presentación debe quedar claramente demostrada la estructura de una célula eucariótica a partir de su descomposición en cada pieza que la conforman, y la explicación de las funciones que cada pieza desempeña. Para efectos prácticos podrán seleccionar una célula vegetal o una animal para elaborar su proyecto.
Siga el siguiente enlace para ir al tutorial de cómo usar PREZI:
https://www.youtube.com/watch?v=9ZI8tsS8mi4
CLASE 3: GRADOS DE ORGANIZACIÓN DE LOS SERES VIVOS
INTRODUCCIÓN
La diversidad de la vida está en el grado de especialidad que han logrado las células y la manera como ellas se integran para conformar un individuo. Muchos organismos son unicelulares microscópicos, otros por su parte son pluricelulares y visibles a nuestros ojos. Una de la principales razones por la cual las células no alcanzan grandes dimensiones es que les resulta ineficiente hacerlo; todos los materiales deben pasar hacia adentro o fuera de la célula a través de la membrana celular, de modo que el tamaño de dicha membrana, en comparación con el del resto de la célula, es un factor crítico. Al aumentar su tamaño un organismo unicelular corre el riesgo de sucumbir a las fuerzas físicas que gobiernan el planeta y por lo tanto perecer si, por ejemplo, los elementos moleculares de su membrana se desintegran. Entre mayor sea el tamaño de una célula esta estará más expuesta a las fuerzas gravitatorias y por lo tanto estará más propensa a sufrir daños mecánicos, es por ello que casi todos los seres unicelulares son microscópicos.
En consecuencia, el aumento de volumen corporal de los organismos vivos ha sido el resultado de la complejisación ocasionada por la unión de cientos, miles y millones de células; gracias a esto aparecieron los organismos coloniales y pluricelulares en el planeta, los cuales evolucionaron, como se esperaría, de los organismos unicelulares. Pero tener un cuerpo con muchas células propone un nuevo reto adaptativo, pues si bien los organismos unicelulares realizan casi todas sus funciones vitales mediante procesos difusivos, los organismos pluricelulares debieron implementar estrategias evolutivas anatómicas que permitieran la nutrición, la respiración, la excreción, entre otras actividades, a cada una de las células que los integran. Debido a esto surgen en los organismos pluricelulares una división celular que dio como resultado el surgimiento de tejidos y órganos especializados en realizar todas estas funciones, de tal manera que que dichos organismos (hongos, plantas y animales) se adaptaran favorablemente a su ambiente.
La organización anatómica de los organismos vivos está dividida en niveles que van del más incipiente al más complejo:
1. Nivel químico:
Es el nivel de organización más simple. Este nivel abarca las partículas básicas de toda la materia, los átomos, y sus combinaciones, llamadas moléculas. Un átomo es la unidad más pequeña de un elemento químico (sustancia fundamental) que aun conserva las propiedades características de dicho elemento . Por ejemplo, un átomo de hierro es la cantidad más pequeña que puede obtenerse de ese elemento. Los átomos se combinan químicamente para conformar las moléculas, por ejemplo, dos átomos de hidrógeno se combinan con uno de oxígeno para formar agua.
1. Nivel celular:
Se corresponde de lleno, tal como dice uno de los postulados de la teoría celular, con las características de un ser vivo. Este nivel está representado por las células que integran al individuo. Se puede decir que una célula es:
- Una unidad vital: es el ser vivo más pequeño que puede existir
- Una unida anatómica: seres vivos constituidos por una o más células
- Una unidad fisiológica: con mecanismos biológicos para sus funciones vitales
- Una unidad genética: deriva de otras preexistentes y transmite la herencia
Cuando las células, como seres individuales, deciden reunirse organizadamente para adaptarse mejor al medio, sin perder su individualidad (cada una de ellas sigue realizando todas y cada una de sus funciones), decimos que las células forman colonias. Entonces, Una colonia celular es un grupo de células con similares características, que actúan en conjunto, con la particularidad de no formar una unidad estructural mayor o tejido. Esta organización se observa tanto en organismos unicelulares procariotas como eucariotas. Ejemplo de esta agregación son las que realizan algas verdes unicelulares como volvox, eudorina, pandorina, etc.

Si esta unión termina con la individualidad de cada una de ellas en busca del bien común, dependiendo unas de otras, entonces hablamos de estructura talófita (típica de hongos, algunas algas eucariotas y plantas talófitas no vasculares como la hepática de la imágen). No se llegan a formar tejidos especializados pero la agrupación comparte sus metabolitos, nutrientes,...haciéndolas depender unas de otras.
Cuando por fin las células unidas se especializan tanto que cada grupo realiza determinadas funciones y no otras, entonces hablamos de tejidos. Los tejidos serían un conjunto de células con el mismo origen embriológico, y con la misma función dentro de un organismo.
La organización de varios tejidos diferentes para desarrollar funciones concretas se conoce como órgano (por ejemplo el corazón).
La agrupación de varios órganos para desarrollar una función biológica se conoce como aparato (por ejemplo el aparato digestivo).
Un sistema es una agrupación de órganos para realizar funciones dispares y distintas (Sistema Nervioso).
Individuo: una o más células caracterizadas por un único tipo de información en su ADN. Puede ser unicelular o pluricelular. Los individuos pluricelulares muestran tipos celulares especializados y división de funciones en tejidos, órganos y sistemas.
3. Nivel ecológico:
Los organismos interactúan entre sí para formar niveles de organización biológica aún más complejos.
Especie: grupo de individuos similares que tienden a aparearse entre sí dando origen a una cría fértil. Todos los organismos de una misma especie que habita en la misma área geográfica conforman una población.
Comunidad: es la relación entre poblaciones de diferentes especies. La estructura de una comunidad puede verse alterada debido por ejemplo al fuego, actividad human o la sobre población.
Ecosistema: el conjunto de seres vivos que se desarrollan con otros individuos de otras especies y con su medio ambiente. Se llama biotopo al lugar donde ocurre esta relación y biocenosis al conjunto de poblaciones interrelacionadas. Así el biotopo y la biocenosis forman el ecosistema.

ACTIVIDAD
1. Reúnase con su equipo de trabajo y elabore un plegable o folleto en el que se describa un grado de organización de los seres vivos (solo uno, el que seleccione el equipo).
2. El diseño del folleto elaborado deberá incluir una primera sección de presentación (institución, programa, asignatura, profesor y participantes). Seguidamente en el folleto se deberá abordar el desarrollo del tema, es decir, organizar el contenido temático teniendo en cuenta la relación de las características estructurales, anatómicas y/o morfológicas principales del grado de organización seleccionado (apóyese en la información de arriba). El estilo de edición del folleto es libre!
No hay comentarios:
Publicar un comentario